First-in-human study of epidural spinal cord stimulation in individuals with spinal muscular atrophy – Nature.com
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain
the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in
Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles
and JavaScript.Advertisement
Nature Medicine
(2025)Cite this article
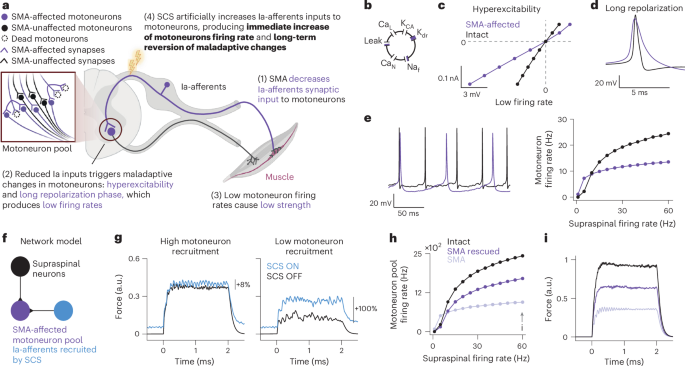
320 Accesses676 AltmetricMetrics detailsSpinal muscular atrophy (SMA) is an inherited neurodegenerative disease causing motoneuron dysfunction, muscle weakness, fatigue and early mortality. Three new therapies can slow disease progression, enabling people to survive albeit with lingering motor impairments. Indeed, weakness and fatigue are still among patients’ main concerns. Here we show that epidural spinal cord stimulation (SCS) improved motoneuron function, thereby increasing strength, endurance and gait quality, in three adults with type 3 SMA. Preclinical works demonstrated that SMA motoneurons show low firing rates because of a loss of excitatory input from primary sensory afferents. In the present study, we hypothesized that correcting this loss with electrical stimulation of the sensory afferents could improve motoneuron function. To test this hypothesis, we implanted three adults with SMA with epidural electrodes over the lumbosacral spinal cord, targeting sensory axons of the legs. We delivered SCS for 4 weeks, 2 h per day during motor tasks. Our intervention led to improvements in strength (up to +180%), gait quality (mean step length: +40%) and endurance (mean change in 6-minute walk test: +26 m), paralleled by increased motoneuron firing rates. These changes persisted even when SCS was turned OFF. Notably, no adverse events related to the stimulation were reported. ClinicalTrials.gov identifier: NCT05430113.This is a preview of subscription content, access via your institution
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
24,99 € / 30 days
cancel any timeSubscribe to this journal
Receive 12 print issues and online access
We are sorry, but there is no personal subscription option available for your country.Buy this articlePrices may be subject to local taxes which are calculated during checkoutThe data that support the findings of this study are openly available at https://doi.org/10.5281/zenodo.14201208. To protect confidentiality of participants, raw data requests should be made officially to M.C. or R.M.F. Reasonable requests for data will be fulfilled upon IRB approval via official material transfer agreements between the University of Pittsburgh and the requesting party, taking into account any existing sponsored research contract agreements with the funders.The code of the biophysical model can be found at https://github.com/genisprat/BiophysicalModel_SCSinSMA.Yasuda, T., Nakata, Y. & Mochizuki, H. α-Synuclein and neuronal cell death. Mol. Neurobiol. 47, 466–483 (2013).Article
CAS
PubMed
Google Scholar
Saxena, S. & Caroni, P. Selective neuronal vulnerability in neurodegenerative diseases: from stressor thresholds to degeneration. Neuron 71, 35–48 (2011).Article
CAS
PubMed
Google Scholar
Blesa, J. & Przedborski, S. Parkinson’s disease: animal models and dopaminergic cell vulnerability. Front. Neuroanat. 8, 155 (2014).Article
PubMed
PubMed Central
Google Scholar
Friedlander, R. M., Brown, R. H., Gagliardini, V., Wang, J. & Yuan, J. Inhibition of ICE slows ALS in mice. Nature 388, 31–31 (1997).Article
CAS
PubMed
Google Scholar
Ona, V. O. et al. Inhibition of caspase-1 slows disease progression in a mouse model of Huntington’s disease. Nature 399, 263–267 (1999).Article
CAS
PubMed
Google Scholar
Li, M. et al. Functional role of caspase-1 and caspase-3 in an ALS transgenic mouse model. Science 288, 335–339 (2000).Article
CAS
PubMed
Google Scholar
Friedlander, R. M. Apoptosis and caspases in neurodegenerative diseases. N. Engl. J. Med. 348, 1365–1375 (2003).Article
CAS
PubMed
Google Scholar
Palop, J. J. & Mucke, L. Network abnormalities and interneuron dysfunction in Alzheimer disease. Nat. Rev. Neurosci. 17, 777–792 (2016).Article
CAS
PubMed
PubMed Central
Google Scholar
Palop, J. J. & Mucke, L. Amyloid-β–induced neuronal dysfunction in Alzheimer’s disease: from synapses toward neural networks. Nat. Neurosci. 13, 812–818 (2010).Article
CAS
PubMed
PubMed Central
Google Scholar
Babiloni, C. et al. What electrophysiology tells us about Alzheimer’s disease: a window into the synchronization and connectivity of brain neurons. Neurobiol. Aging 85, 58–73 (2020).Article
PubMed
Google Scholar
Held, A. et al. Circuit dysfunction in SOD1-ALS model first detected in sensory feedback prior to motor neuron degeneration is alleviated by BMP signaling. J. Neurosci. 39, 2347–2364 (2019).Article
CAS
PubMed
PubMed Central
Google Scholar
Mentis, G. Z. et al. Early functional impairment of sensory-motor connectivity in a mouse model of spinal muscular atrophy. Neuron 69, 453–467 (2011).Article
CAS
PubMed
PubMed Central
Google Scholar
Mercuri, E. et al. Safety and efficacy of once-daily risdiplam in type 2 and non-ambulant type 3 spinal muscular atrophy (SUNFISH part 2): a phase 3, double-blind, randomised, placebo-controlled trial. Lancet Neurol. 21, 42–52 (2022).Article
CAS
PubMed
Google Scholar
Tisdale, S. & Pellizzoni, L. Disease mechanisms and therapeutic approaches in spinal muscular atrophy. J. Neurosci. 35, 8691–8700 (2015).Article
CAS
PubMed
PubMed Central
Google Scholar
Lefebvre, S. et al. Identification and characterization of a spinal muscular atrophy-determining gene. Cell 80, 155–165 (1995).Article
CAS
PubMed
Google Scholar
Arnold, W. D., Kassar, D. & Kissel, J. T. Spinal muscular atrophy: diagnosis and management in a new therapeutic era. Muscle Nerve 51, 157–167 (2015).Article
CAS
PubMed
Google Scholar
Reilly, A., Chehade, L. & Kothary, R. Curing SMA: are we there yet? Gene Ther. 30, 8–17 (2023).Article
CAS
PubMed
Google Scholar
Fletcher, E. V. et al. Reduced sensory synaptic excitation impairs motor neuron function via Kv2. 1 in spinal muscular atrophy. Nat. Neurosci. 20, 905–916 (2017).Article
CAS
PubMed
PubMed Central
Google Scholar
Alonso, L. M., Rue, M. C. & Marder, E. Gating of homeostatic regulation of intrinsic excitability produces cryptic long-term storage of prior perturbations. Proc. Natl Acad. Sci. USA 120, e2222016120 (2023).Article
PubMed
PubMed Central
Google Scholar
Simon, C. M. et al. Chronic pharmacological increase of neuronal activity improves sensory-motor dysfunction in spinal muscular atrophy mice. J. Neurosci. 41, 376–389 (2021).Article
CAS
PubMed
PubMed Central
Google Scholar
Blackburn, A. Z. et al. Spinal cord stimulation via percutaneous and open implantation: systematic review and meta-analysis examining complication rates. World Neurosurg. 154, 132–143 (2021).Article
PubMed
Google Scholar
Capogrosso, M. et al. A computational model for epidural electrical stimulation of spinal sensorimotor circuits. J. Neurosci. 33, 19326–19340 (2013).Article
CAS
PubMed
PubMed Central
Google Scholar
Rattay, F., Minassian, K. & Dimitrijevic, M. R. Epidural electrical stimulation of posterior structures of the human lumbosacral cord: 2. quantitative analysis by computer modeling. Spinal Cord 38, 473–489 (2000).Article
CAS
PubMed
Google Scholar
McIntyre, C. C. & Grill, W. M. Extracellular stimulation of central neurons: influence of stimulus waveform and frequency on neuronal output. J. Neurophysiol. 88, 1592–1604 (2002).Article
PubMed
Google Scholar
Formento, E. et al. Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury. Nat. Neurosci. 21, 1728–1741 (2018).Article
PubMed
PubMed Central
Google Scholar
Moraud, E. M. et al. Mechanisms underlying the neuromodulation of spinal circuits for correcting gait and balance deficits after spinal cord injury. Neuron 89, 814–828 (2016).Article
CAS
PubMed
Google Scholar
Balaguer, J.-M. et al. Supraspinal control of motoneurons after paralysis enabled by spinal cord stimulation. Preprint at medRxiv https://doi.org/10.1101/2023.11.29.23298779 (2023).Booth, V., Rinzel, J. & Kiehn, O. Compartmental model of vertebrate motoneurons for Ca2+-dependent spiking and plateau potentials under pharmacological treatment. J. Neurophysiol. 78, 3371–3385 (1997).Article
CAS
PubMed
Google Scholar
Wagner, F. B. et al. Targeted neurotechnology restores walking in humans with spinal cord injury. Nature 563, 65 (2018).Article
CAS
PubMed
Google Scholar
Powell, M. P. et al. Epidural stimulation of the cervical spinal cord for post-stroke upper-limb paresis. Nat. Med. 29, 689–699 (2023).Article
CAS
PubMed
PubMed Central
Google Scholar
Kinany, N. et al. Towards reliable spinal cord fMRI: assessment of common imaging protocols. Neuroimage 250, 118964 (2022).Article
PubMed
Google Scholar
Capogrosso, M. et al. Configuration of electrical spinal cord stimulation through real-time processing of gait kinematics. Nat. Protoc. 13, 2031–2061 (2018).Article
CAS
PubMed
Google Scholar
Rowald, A. et al. Activity-dependent spinal cord neuromodulation rapidly restores trunk and leg motor functions after complete paralysis. Nat. Med. 28, 260–271 (2022).Article
CAS
PubMed
Google Scholar
Greiner, N. et al. Recruitment of upper-limb motoneurons with epidural electrical stimulation of the primate cervical spinal cord. Nat. Commun. 12, 435 (2021).Article
CAS
PubMed
PubMed Central
Google Scholar
Sharpe, A. N. & Jackson, A. Upper-limb muscle responses to epidural, subdural and intraspinal stimulation of the cervical spinal cord. J. Neural Eng. 11, 016005 (2014).Article
PubMed
PubMed Central
Google Scholar
Hofstoetter, U. S. et al. Periodic modulation of repetitively elicited monosynaptic reflexes of the human lumbosacral spinal cord. J. Neurophysiol. 114, 400–410 (2015).Article
CAS
PubMed
PubMed Central
Google Scholar
Hultborn, H. et al. On the mechanism of the post-activation depression of the H-reflex in human subjects. Exp. Brain Res. 108, 450–462 (1996).Article
CAS
PubMed
Google Scholar
Šarabon, N., Kozinc, Ž. & Perman, M. Establishing reference values for isometric knee extension and flexion strength. Front. Physiol. 12, 767941 (2021).Article
PubMed
PubMed Central
Google Scholar
Bartels, B., Montes, J., van der Pol, W. L. & de Groot, J. F. Physical exercise training for type 3 spinal muscular atrophy. Cochrane Database Syst. Rev. 3, CD012120 (2019).PubMed
Google Scholar
Mercuri, E., Sumner, C. J., Muntoni, F., Darras, B. T. & Finkel, R. S. Spinal muscular atrophy. Nat. Rev. Dis. Prim. 8, 52 (2022).Article
PubMed
Google Scholar
Montes, J. et al. A randomized, controlled clinical trial of exercise in patients with spinal muscular atrophy: methods and baseline characteristics. J. Neuromuscul. Dis. 1, 151–161 (2014).Article
PubMed
Google Scholar
Montes, J. et al. Single-blind, randomized, controlled clinical trial of exercise in ambulatory spinal muscular atrophy: why are the results negative? J. Neuromuscul. Dis. 2, 463–470 (2015).Article
PubMed
PubMed Central
Google Scholar
Montes, J. et al. Six-Minute Walk Test demonstrates motor fatigue in spinal muscular atrophy. Neurology 74, 833–838 (2010).Article
CAS
PubMed
PubMed Central
Google Scholar
Farina, D. et al. Estimation of single motor unit conduction velocity from surface electromyogram signals detected with linear electrode arrays. Med. Biol. Eng. Comput. 39, 225–236 (2001).Article
CAS
PubMed
Google Scholar
Del Vecchio, A. et al. You are as fast as your motor neurons: speed of recruitment and maximal discharge of motor neurons determine the maximal rate of force development in humans. J. Physiol. 597, 2445–2456 (2019).Article
PubMed
PubMed Central
Google Scholar
Van Cutsem, M., Duchateau, J. & Hainaut, K. Changes in single motor unit behaviour contribute to the increase in contraction speed after dynamic training in humans. J. Physiol. 513, 295–305 (1998).Article
PubMed
PubMed Central
Google Scholar
Del Vecchio, A., Falla, D., Felici, F. & Farina, D. The relative strength of common synaptic input to motor neurons is not a determinant of the maximal rate of force development in humans. J. Appl. Physiol. 127, 205–214 (2019).Article
PubMed
Google Scholar
Norton, J. A., Roy, F. D. & Mahood, J. K. Preservation of motor evoked potentials under anesthesia in children with spinal muscular atrophy type II undergoing spinal deformity surgery. J. Clin. Neurophysiol. 30, 382–385 (2013).Article
PubMed
Google Scholar
Jiang, Y.-Q., Zaaimi, B. & Martin, J. H. Competition with primary sensory afferents drives remodeling of corticospinal axons in mature spinal motor circuits. J. Neurosci. 36, 193–203 (2016).Article
CAS
PubMed
PubMed Central
Google Scholar
Türker, K. S. & Cheng, H. B. Motor-unit firing frequency can be used for the estimation of synaptic potentials in human motoneurones. J. Neurosci. Methods 53, 225–234 (1994).Article
PubMed
Google Scholar
Metz, K. et al. Post‐activation depression from primary afferent depolarization (PAD) produces extensor H‐reflex suppression following flexor afferent conditioning. J. Physiol. 601, 1925–1956 (2023).Article
CAS
PubMed
Google Scholar
Borg, J. Refractory period of single motor nerve fibres in man. J. Neurol. Neurosurg. Psychiatry 47, 344–348 (1984).Article
CAS
PubMed
PubMed Central
Google Scholar
Simon, C. M. et al. Dysfunction of proprioceptive sensory synapses is a pathogenic event and therapeutic target in mice and humans with spinal muscular atrophy. Preprint at medRxiv https://doi.org/10.1101/2024.06.03.24308132 (2024).Arslan, D. et al. Nusinersen for adults with spinal muscular atrophy. Neurol. Sci. 44, 2393–2400 (2023).Article
PubMed
Google Scholar
Angeli, C. A. et al. Recovery of over-ground walking after chronic motor complete spinal cord injury. N. Engl. J. Med. 379, 1244–1250 (2018).Article
PubMed
Google Scholar
Gill, M. L. et al. Neuromodulation of lumbosacral spinal networks enables independent stepping after complete paraplegia. Nat. Med. 24, 1677–1682 (2018).Article
CAS
PubMed
Google Scholar
Squair, J. W. et al. Implanted system for orthostatic hypotension in multiple-system atrophy. N. Engl. J. Med. 386, 1339–1344 (2022).Article
PubMed
Google Scholar
Milekovic, T. et al. A spinal cord neuroprosthesis for locomotor deficits due to Parkinson’s disease. Nat. Med. 29, 2854–2865 (2023).Article
CAS
PubMed
Google Scholar
Hofstoetter, U. S., Freundl, B., Lackner, P. & Binder, H. Transcutaneous spinal cord stimulation enhances walking performance and reduces spasticity in individuals with multiple sclerosis. Brain Sci. 11, 472 (2021).Article
PubMed
PubMed Central
Google Scholar
Holobar, A. & Zazula, D. Multichannel blind source separation using convolution kernel compensation. IEEE Trans. Signal Process. 55, 4487–4496 (2007).Article
Google Scholar
Del Vecchio, A. et al. Tutorial: analysis of motor unit discharge characteristics from high-density surface EMG signals. J. Electromyogr. Kinesiol. 53, 102426 (2020).Article
PubMed
Google Scholar
Hug, F. et al. Analysis of motor unit spike trains estimated from high-density surface electromyography is highly reliable across operators. J. Electromyogr. Kinesiol. 58, 102548 (2021).Article
PubMed
Google Scholar
Holobar, A., Minetto, M. A. & Farina, D. Accurate identification of motor unit discharge patterns from high-density surface EMG and validation with a novel signal-based performance metric. J. Neural Eng. 11, 016008 (2014).Article
CAS
PubMed
Google Scholar
Frančič, A. & Holobar, A. On the reuse of motor unit filters in high density surface electromyograms with different signal-to-noise ratios. In 8th European Medical and Biological Engineering Conference (eds Jarm, T. et al.) 923–931 (Springer International, 2021).Download referencesThis study was supported by an exploratory research grant from F. Hoffmann-La Roche to M.C. and R.M.F. S.D. is supported by a joint research grant from the SMA Foundation and SMA Europe. MRI imaging was supported by Carnegie Mellon University, CMU-Pitt BRIDGE Center, RRID:SCR_023356. We would like to thank S. Bader for the support as clinical coordinator for this work. We thank the participants of this study for being pioneers in neurostimulation research and for the great insight into the real needs of people living with SMA. We thank B. Petersen for helping set up the experiments in SMA01. We also thank L. Pellizzoni for useful conversations on the mechanisms of SMA. Finally, we thank the SMA Foundation and specifically L. Eng for advice, support and encouragement.These authors contributed equally: Genís Prat-Ortega, Scott Ensel, Serena Donadio.These authors jointly supervised this work: Elivra Pirondini, Robert M. Friedlander and Marco Capogrosso.Department of Neurological Surgery, University of Pittsburgh, Pittsburgh, PA, USAGenís Prat-Ortega, Serena Donadio, Jonathan Ho, Sarah Frazier-Kim, Daryl P. Fields, Jeffrey Balzer, Peter Gerszten, Robert M. Friedlander & Marco CapogrossoRehab Neural Engineering Labs, University of Pittsburgh, Pittsburgh, PA, USAGenís Prat-Ortega, Scott Ensel, Serena Donadio, Amy Boos, Jonathan Ho, Erick Carranza, Sarah Frazier-Kim, Daryl P. Fields, Lee E. Fisher, Elvira Pirondini & Marco CapogrossoDepartment of Bioengineering, University of Pittsburgh, Pittsburgh, PA, USAScott Ensel, Serena Donadio, Erick Carranza, Lee E. Fisher, Elvira Pirondini & Marco CapogrossoDepartment of Mechanical engineering, Carnegie Mellon University, Pittsburgh, PA, USALuigi Borda, Prakarsh Yadav, Nikhil Verma & Doug J. WeberNeuroMechatronics Lab, Carnegie Mellon University, Pittsburgh, PA, USALuigi Borda, Prakarsh Yadav, Nikhil Verma & Doug J. WeberDepartment of Neurology, University of Pittsburgh, Pittsburgh, PA, USAAmy Boos & Paula R. ClemensDepartment of Physical Medicine and Rehabilitation, University of Pittsburgh, Pittsburgh, PA, USALee E. Fisher & Elvira PirondiniDepartment of Biomedical Engineering, Carnegie Mellon University, Pittsburgh, PA, USALee E. FisherThe Neuroscience Institute, Carnegie Mellon University, Pittsburgh, PA, USADoug J. WeberDepartment of Neurology and Clinical Neurosciences, Stanford University, Palo Alto, CA, USATina DuongGenentech Roche, San Francisco, CA, USASteven D. Weinstein & Mikael J. L. EliassonDepartment of Rehabilitation and Regenerative Medicine, Columbia University, New York, NY, USAJacqueline MontesSpinal Muscular Atrophy Foundation New York, New York, NY, USAKaren S. ChenCenter for Motor Neuron Biology and Disease, Columbia University, New York, NY, USAGeorge Z. MentisDepartment of Neurology, Columbia University, New York, NY, USAGeorge Z. MentisDepartment of Pathology and Cell Biology, Columbia University, New York, NY, USAGeorge Z. MentisClinical and Translational Science Institute (CTSI), University of Pittsburgh, Pittsburgh, PA, USAElvira PirondiniYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarYou can also search for this author in
PubMed Google ScholarM.C., R.M.F. and M.J.L.E. conceived the study. G.P.O., S.E. and S.D. performed and designed all experiments and analyzed the data. L.B. and N.V. performed single motoneuron experiments. G.P.O., S.E., S.D. and L.B. created figures. L.B., P.Y., N.V. and D.W. analyzed single motoneuron data. A.B. performed all clinical evaluations, under the supervision of T.D., J.M. and P.C. S.F.-K. analyzed physical activity data. D.P.F. and P.G. implanted all participants and designed implant procedures, with M.C. and L.E.F. M.J.L.E., S.D.W., K.S.C., M.C. and T.D. designed the clinical study. P.C. directed participant assessments and eligibility. J.H. performed and analyzed TMS data. E.C. performed proprioception experiments. S.E. and E.P. conceived and performed all fMRI experiments and analysis. J.B., M.C. and L.E.F. designed intraoperative electrophysiology procedures for lead placement. J.M. provided the control dataset and supported the interpretation of clinical outcomes. G.P.O. and M.C. conceived and implemented the computational model. M.C., E.P., R.M.F. and G.Z.M. directed the scientific interpretation of data. R.M.F., E.P. and M.C. directed all experimental activities. R.M.F. and M.C. secured funding for the study. G.P.O., S.E., S.D., E.P., M.C. and R.M.F. wrote the paper, and all authors contributed to its editing.Correspondence to
Elvira Pirondini, Robert M. Friedlander or Marco Capogrosso.This study was supported by an exploratory research grant from F. Hoffmann-La Roche to M.C., and Genentech and the University of Pittsburgh hold rights to intellectual property related to this study. M.C., G.P.O. and M.J.L.E. hold patent applications that relate to this work. The remaining authors declare no competing interests.Nature Medicine thanks the anonymous reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Jerome Staal, in collaboration with the Nature Medicine team.Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.a-b Rheobase for WT and SMA motoneuron biophysical models. To compute the rheobase we slowly increased the input current injected to the motoneuron’s soma (a) until a spike was triggered (b). The black and purple lines indicated the rheobase, the smallest amplitude that triggers an action potential. The SMA motoneuron biophysical model reproduces the active hyperexcitability characterized by reduced rheobase. c, Example of 2 second isometric force produced by a SMA-affected motoneuron pool simulated with the biophysical model. From bottom to top: Raster plot of 11 Ia-afferent fibers recruited by SCS at 40 Hz. Note that spikes are completely synchronized because they are triggered by SCS pulses. Supraspinal neurons with natural firing rate (Poisson neurons); for visualization propose, we only show 11 neurons but the supraspinal population has 200 neurons. Motoneuron pool firing rate with 30% of motoneuron loss and 50% SMA-affected motoneurons. Simulated force produced by the motoneuron pool normalized by the maximum force produced by an intact motoneuron pool. d, SMA-affected motoneuron pool firing rate for different simulated percentage of motoneuron loss and SMA-affected motoneurons. e, Simulated force for different percentages of motoneuron loss and SMA-affected motoneurons. f, Motoneuron pool firing rate as a function of the supraspinal firing rate with SCS ON and OFF. The biophysical model predicted that the effect of SCS should be more prominent on low than high supraspinal inputs indicating that if all motoneurons are recruited at their maximum firing rate, there should be no effect of SCS. Thus, the effect of SCS at maximum voluntary contraction will depend on the participants’ ability to recruit the resources of their motoneuron pool. SCS could give access to motoneurons that are not recruited or recruited with a submaximal firing rate. g, Simulated force with a low supraspinal input (5 Hz) and very high supraspinal input (50 Hz). h, Firing rate for a SMA-affected pool with all SMA-affected motoneurons and 40% rescued which is the percentage of rescued motoneurons for SMA01 Right Knee Extension. i, Predicted forces by the biophysical model given the percentage of motoneurons rescued in SMA01 Right Knee Extension and SMA03 Right Knee Flexion. First, we simulated the torque produced with a SMA-affected pool with 70 motoneurons all affected by SMA (light purple). In purple we simulated the torque rescuing the same percentage that we found experimentally: 40% for SMA01 Right Knee Extension and 25% SMA03 Right Knee Flexion. Finally, we used a bootstrap method to generate multiple predictions of the percentage of rescued units per participant, to estimate a maximum possible upper limit that could occur by random variations in the data (95th percentile). Using this method we found that the maximum percentages of rescued motoneuron consistent with our data were 80% and 50% for SMA01 Right Knee Extension and SMA03 Right Knee Flexion respectively. We simulated the torques with these upper limits (dark purple) and we found that the experimental values (dash black line) fell below the upper boundary. For SMA01 our biophysical model predicted an increase of 24% and up to 69% which is consistent with the experimental increase in right knee extension +55%. For SMA03 our model predicted an increase in torque of 15% and up to 42% which is remarkably consistent with the +21% increase that we found in right knee flexion. All simulated torques are normalized by the torque produced by a SMA-affected motoneuron pool with 70 SMA-motoneurons. j, Simulated peak to peak of the TMS MEP for two hypotheses: reversion of the maladaptive changes produced by SMA (SMA rescued) and reduction of the supraspinal synaptic weight. k, Examples of simulated MEP. Both hypotheses are consistent with experimental findings of smaller MEP. l,m, Simulated firing rates as a function of the supraspinal inputs (l) and simulated torque (m) for the two hypotheses. Only the hypothesis of rescued motoneurons is consistent with the experimental findings of higher firing rates and torques.a, d, g, For each study participant demographic information, including ethnicity, age, score at the enrollment on the Hammersmith Functional Motor Scale Expanded (HFMSE), and detail on any treatment used. b, e, h. Left: pre implant T2 MRI, with the approximate lead location in red. Right: X-ray post-implant, with the bilateral leads in black. All three participants had the conus medullaris under the vertebra T12, a vertebra above what is normally observed (L1) c, f, i, SCS parameters used during walking. We used a program targeting the right and a program targeting the left leg muscles. Each program applied a fixed amplitude, frequency and pulse width for each SCS pulse. j, l, n. Configurations of the experimental set ups for passive knee joint movement for all participants and the SCS parameters used (anode blue, cathode red). k, m, o. Muscle responses were used to compute the peak to peak of each bin. Plots are reporting the mean and 95% CI of the peak-to-peak responses. All error bars indicate the confidence interval, computed with bootstrap (N = 10,000). p. The cycle of joint oscillations was divided into 10 bins of equal duration during which the muscle responses were extracted and regrouped. q. Configuration of the experimental set up for proprioception testing (Supplementary Methods). Randomly selected flexion or extension movement were imposed to the knee joint of each subject at a speed of 0.5 degree per second. r. Configurations of SCS parameters used (anode blue, cathode red) and the detection angle (the difference between the starting angle and final angle displacement) plotted for each participant under varying SCS conditions of the proprioception testing: SCS OFF (black), SCS ON (light blue). The SCS ON condition was continuous stimulation at 40 Hz. Statistical differences were assessed with two-sided bootstrap (N = 10,000). *, **, and *** denote significant differences with p-values of p < 0.05, p < 0.01, and p < 0.001, respectively.a,Maximum strength in hip and knee for all participants at pre-implant, post-explant and follow up (6 weeks after post explant). To reduce the day-to-day variability in our measures of maximum torque, we used data from all session’s pre-implant and post-explant (see Extended Data Fig. 4b–f). We corrected for multiple comparison using Bonferroni with N = 3 (pre-implant vs post-explant, pre-implant vs follows up and post-explant vs follow up) b, Maximum strength with SCS ON and OFF for all sessions normalized by the mean maximum torque with SCS OFF across repetitions for of each session. Error bars show the mean and 95% confidence interval of the mean, computed with bootstrap (N = 10,000). Each dot represents the torque of a single repetition. All statistical significance was assessed with two-sided bootstrapping (N = 10,000): p < 0.05 (*), p < 0.01 (**), p < 0.001(***).a, Experimental set up to measure isometric torques for knee extension and flexion. b-f Maximum voluntary contraction in all muscles, sessions and subjects that we could reliably measure (torque above 4 N): SMA01 Hip (b), SMA01 Knee (c), SMA02 Hip (d), SMA03 Hip (e) and SMA03 (f). Each point corresponds to one repetition and the square represents the mean across repetitions. g, We used a multivariable linear regression analysis to study the effect of fatigue in our measurements of torque (see Methods). For SMA01, we found that the largest regressor was the repetition indicating that SMA01 was fatiguing after only a few repetitions of maximum voluntary contractions. Given that we always started with SCS OFF trials, our results could be underestimating the assistive effect of SCS (Fig. 3a-c). Indeed, we found for SMA01 right knee extension, the interaction between the SCS and the repetition number was significant indicating that SCS increased maximum torque when SMA01 was fatigued. In SMA03 we reduced the trial duration time from 5 to 2 seconds and we found that the repetition number had no impact indicating that direct comparison between SCS OFF and ON trial is probably not affected by the repetition number. h,Torque traces for SCS ON and SCS OFF fatigued repetition (after more than 12 repetitions of maximum voluntary contraction) in day 12 (See supplementary methods for more information). i, Maximum torque at right knee extension produced by SMA01 in fatigued trials for each session. j, Data from all sessions pooled where we tested the maximum torque in fatigued condition. All error bars show the mean and 95% confidence interval of the mean, computed with bootstrap (N = 10,000).a. Gait variables (step length, step height, and stride velocity) for SCS ON and OFF for all sessions normalized by the median value with SCS OFF across repetitions for each session. P-values corrected with Bonferroni correction (N = 2, 4 and 4 for SMA01, SMA02 and SMA03 respectively) b, Same gait variables for each session with SCS OFF. We used Kruskal Wallis ANOVA to find significance differences between groups (all p-values < 0.001). To study if the improvements saturated, we compared each week with the following week. In most gait quality variables, the improvements do not saturate (significance difference between week 3 and week 4). P-values corrected using Bonferroni correction (N = 3, 4 and 4 for SMA01, SMA02 and SMA03 respectively) c, Therapeutics effects in gait quality variables (week 1 vs week 4). All gait quality variables improved after the SCS intervention. d. Left, Examples of isometric torque traces during maximum voluntary knee extension contraction with SCS ON, SCS OFF, and with sham SCS parameters. The sham consisted of an SCS configuration chosen not to target the specific muscles involved in the joint movement tested changing the active contacts or the amplitude or frequency of the stimulation. Right, Maximum torque during knee extension for SCS ON, OFF, and sham SCS parameters. Dots represent individual repetitions, while error bars indicate mean and the 95% CI of the mean (CI). e. Maximum torque during Hip Flexion for SMA01 using the same program with different stimulation amplitudes. Dots represent individual repetitions, while error bars indicate mean and the 95% CI of the mean (CI). f. Left, Mean heel marker trajectory across gait cycles with SCS ON using same program and different frequencies. Right, gait quality variables computed under the same conditions. P-values corrected using Bonferroni correction (N = 7). g, Setup for testing exaggerated hip flexion walking on the anti-gravity treadmill (Supplementary Methods). h. Left, Examples of metatarsal marker trajectory during exaggerated hip flexion walking with SCS ON and OFF. Right, The maximum hip flexion increased with SCS ON. i, same to h, considering SCS OFF in week 1 vs week 4. Again, the maximum hip flexion increased from week 1 to week 4 even without SCS. Box plots represent the median, 25th and 75th percentile and minimum and maximum data points without outliers. All statistical significance was assessed with two-sided bootstrapping (N = 10,000) and corrected for multiple comparisons: p < 0.05 (*), p < 0.01 (**), p < 0.001(***).a. Scheme depicting the placement of the EMG sensor during overground walking (Supplementary Methods). b-c Assistive effects in range of motion (ROM) for SMA03 during week1. b, Top: Mean of hip flexion/extension across gait cycles with SCS OFF (black) and SCS ON (light blue). Bottom: Mean envelope of Semitendinosus (Semi) across the gait cycle with SCS OFF (black) and SCS ON (light blue). The gold shades highlight semitendinosus activation in the pre-swing, contributing to hip extension during the same phase of the cycle. c. Top: Mean change in ROM for the left hip and knee across gait cycles (n = 18, SCS ON; n = 17 SCS OFF). Bottom: Change in EMG activity, computed as the root mean square, for Semitendinosus (Semi), Vastus Lateralis (VastLate), and Biceps Femoris (BicepFem). Improvements in hip ROM should be correlated with the EMG activity at Semi and BicepFem (muscles involved in hip extension). Improvements in knee extension should be related to higher activation on the knee extensors. No change of EMG or ROM. d-e Mean therapeutic improvement in ROM during a gait cycle for SMA03 (n = 17, week 1; n = 16 week 4). Improvements in hip and knee ROM are paralleled by an increase in EMG activity at hip and knee extensors. f-g. Mean assistive improvement in ROM during a gait cycle (n = 60 SCS OFF; n = 28 SCS ON) for SMA02 in week 1. The gray shades highlight Rectus Femoris proximal activation in the swing phase, contributing to hip flexion during the same phase of the cycle. An increase in hip ROM can be explained by an increase in EMG activity at the Rectus Femoris proximal (a proxy of EMG activity of the hip flexors such as the iliopsoas). h-j. Therapeutic improvement in ROM during a gait cycle (n = 60, week 1; n = 18 week 4) for SMA02. Improvements in hip and knee ROM are correlated with increases in EMG activity of hip flexors (rectus femoris proximal) and knee flexors (Semi) No data is reported for SMA01 due to EMG acquisition corruption by noise. Error bars show the mean and 95% confidence interval of the mean, computed with bootstrap (N = 10,000).Comparison with all exercise-only control participants who completed three months of aerobic and strength training, including 3 minors. Changes for the control group were computed by comparing pre- and post- 3 months exercise intervention data, while changes for the SCS group were computed by comparing pre-implant and post-explant data, except otherwise specified. a, Clinical assessments. HFMSE and 6MWT measured pre-implant, post-explant and 6 weeks follow up. In parenthesis changes from baseline measures. All tests executed with SCS OFF. 6MWT dropped back to pre-implant at 6 weeks follow up in 2/3 participants. b, Relationship between age and age of onset for exercise-only control group (black) and the participants in our study (blue). c, Histogram of HFMSE scores for the control group (black), blue lines represent the HFMSE scores for each SCS participant. d, Histogram of aerobic exercise time for the control group (black), and exercise time of each SCS participant (blue lines). e, left: Relationship between the change in 6MWT score and the HFMSE score for the controls (R2 = 0.001) and the SCS participants (R2 = 0.98). Right: Mean and 95%CI of the mean change in 6MWT score for the controls (black) and changes for SCS participants (blue). f, Left: Linear regression analysis between the change in fatigue index during the 6MWT and the severity for both the control group (R2 = 0.15) and the SCS participants (R2 = 0.7). Right: Mean and 95% CI of the mean change in fatigue index across controls (black) and changes for SCS participants (blue) g, left: Regression analysis between the change in stride length and severity for controls (R2 = 0.01), blue dots indicate SCS participants. Right: Mean change in stride length for the controls (black), and changes for SCS participants (blue). h, Same as g for stride velocity (R2 = 0.07). For the SCS participants, changes were computed during self-paced overground walking, comparing SCS OFF trials from week 1 and week 4. i, Left: Regression analysis between the change in knee flexion strength for the controls (R2 = 0.05). Improvements for SMA01 and SMA03 and their confidence intervals are shown in blue. Right: Mean knee flexion strength for the controls (black) and SCS participants (blue). j, Same as i for knee extension (R2 = 0.48). Error bars represent the 95% confidence interval of the mean, computed with bootstrapping (N = 1000).a. Pipeline to extract single motor units spike trains from high density surface electromyography (HDEMG). We used an 8 × 8 channel flexible grid to record muscle activity from both knee extensor (Rectus Femoris (RF)) and flexor (Biceps Femoris (BF)). Participants performed two sets of three isometric Maximum Voluntary Contractions (MVC) of knee extension and flexion. We used the convolution kernel compensation (CKC) method to decompose the EMG signals in single motor units spike trains. b. Rescued motoneuron identification: a motor unit was defined as rescued if its mean peak firing rate (across trials) was higher than the 99.73 % CI (red line) of mean peak firing rate across motor units recorded during the first session. The 99.73% CI was computed with bootstrap (N = 10,000). c. Example of motor units smoothed discharge rate (SDR) obtained during the 3rd MVC. Each line represents the SDR of every motoneuron. Light and dark colors delineate the data from the first and last session where we performed the experiment d. Circles illustrate the number of identified motoneurons over the different phases of the study. (that is, PI = pre-implant; Wi = i-th week after the implant; PE = post-explant). e, Example (SMA02) of the mean and 95% CI of post-explant single motoneurons firing rates during MVC. In gray regions indicate the start and end windows used to compute firing rate at the beginning and the end of the MVC. f, Percentage change in the mean firing rate between rescued and dysfunctional motoneurons at the start (first 150 ms) and end (last 150 ms) of the MVC for each participant. 95% CI were computed using bootstrapping (n = 10,000). We assessed statistical significance using a two-sided test with bootstrapping (n = 10000). Asterisk *, **, and *** denote significant differences with p-values of p < 0.05, p < 0.01, and p< 0.001, respectively.Step 1, Acquisition of functional MRI from the spinal cord in response to the recruitment of motor neurons from specific leg muscles. The motor neurons are recruited by actively extending and flexing the knee joint. Three runs are acquired for each participant. Only the right leg muscles are tested. Instructions were displayed on a screen (fixation cross ‘+’ during rest blocks and text indicating activity blocks), while auditory cues were used to signal changing phases. In addition to the functional volume series, T2 anatomical images and physiological (heart rate, respiratory) signals are acquired. Step 2, Raw fMRI volume series are repeatedly acquired every 2.5 s (TR) in functional runs lasting about 6 minutes. Step 3, Spinal segments are identified from high-definition T2-ZOOMit structural images, T2 images and SCS recruitment curve responses. Spinal segments are then reported in the T2 anatomical image acquired in each run. Step 4, Then motion correction is applied to each functional run in two stages. First, volumes are aligned to the averaged images using 3D rigid body realignment. Second, a slice-by-slice 2D realignment is applied to each volume. Step 5, The spinal cord (including gray and white matter) is automatically segmented from the motion corrected mean functional image, and then manually corrected. Step 6, Functional images from runs two and three are aligned to functional images of run one using the motion corrected mean images by landmarking the conus medullaris and any visible vertebra, using rigid transformations. Step 7, The motion corrected mean images are then coregistered to the PAM50 template using the conus medullaris landmark to extract the transformation matrices, using non-rigid transformations. Step 8, Physiological signals (heart rate and respiratory) acquired concomitantly to the fMRI volumes are used to model physiological noise (RETROICOR based procedure). Acquisition timings corresponding to motion corrected fMRI volume series and physiological noise regressors are submitted to a specific first level generalized linear model (GLM). Residuals from this GLM are then spatially smoothed, volume by volume with 3D gaussian kernel with full width at half maximum (FWHM) of 2x2x6mm3. Acquisition timings corresponding to the task-design and residuals resulting from noise removal are submitted to a specific first level GLM. A second level fixed effects analysis (subject level, task specific) is performed by combining the three runs. Activity pattern maps are uncorrected and thresholded (Z > 2.3, p < 0.01). Step 9, Transformations matrices found in step 7 are then applied to the resulting activity pattern maps of step 8 to arrive in PAM50 space. Step 10, Activity patterns from the three participants, are then compared and analyzed across scanning sessions using the spinal segments found in step 3.a, Quantification of percent change in area under the curve of peak-to-peak TMS responses post-explant vs six week follow up across all intensities tested. Abbreviations: ST, Semitendinosus; BF, Bicep Femoris; VL, Vastus Lateralis; RP, Rectus Femoris Proximal; RD, Rectus Femoris Distal; G, Gastrocnemius; TA, Tibialis Anterior. Error bars are the mean and 95% CI of the mean peak-to-peak responses (bootstrap N = 10,000). b, The SCS parameters used and the resulting plots representing the normalized peak-to-peak response as a function of SCS amplitude. Shaded areas correspond to standard error across responses, while thick lines are the means across responses.SMA01 standing from a kneeling position with and without stimulation.Sit-to-stand transitions with and without stimulation for SMA01.Sit-to-stand transitions with and without stimulation for SMA03.SMA01 exacerbated hip flexion during weight-supported treadmill walking with and without stimulation.SMA02 walking overground with and without stimulation in week 1 and without stimulation in week 4.First lap of the 6MWT of SMA02.Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.Reprints and permissionsPrat-Ortega, G., Ensel, S., Donadio, S. et al. First-in-human study of epidural spinal cord stimulation in individuals with spinal muscular atrophy.
Nat Med (2025). https://doi.org/10.1038/s41591-024-03484-8Download citationReceived: 01 October 2024Accepted: 20 December 2024Published: 05 February 2025DOI: https://doi.org/10.1038/s41591-024-03484-8Anyone you share the following link with will be able to read this content:Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
Advertisement
Nature Medicine (Nat Med)
ISSN 1546-170X (online)
ISSN 1078-8956 (print)
© 2025 Springer Nature LimitedSign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.
Source: https://www.nature.com/articles/s41591-024-03484-8






